Sus scrofa 


System : Terrestrial
Kingdom
Phylum
Class
Order
Family
Animalia
Chordata
Mammalia
Artiodactyla
Suidae
- General
- Distribution
- Impact
- Management
- Bibliography
- Contact

")

Common name
pig (English), Wildschwein (German), razorback (English), te poaka (Maori), kuhukuhu (Maori), poretere (Maori), petapeta (Maori), kune-kune (Maori, New Zealand)
Synonym
Similar species
Summary
Sus scrofa (feral pigs) are escaped or released domestic animals which have been introduced to many parts of the world. They damage crops, stock and property, and transmit many diseases such as Leptospirosis and Foot and Mouth disease. Rooting pigs dig up large areas of native vegetation and spread weeds, disrupting ecological processes such as succession and species composition. Sus scrofa are omnivorous and their diet can include juvenile land tortoises, sea turtles, sea birds, endemic reptiles and macro-invertebrates. Management of Sus scrofa is complicated by the fact that complete eradication is often not acceptable to communities that value feral pigs for hunting and food.
Species Description
Pigs are large omnivorous mammals with powerful bodies and coarse hairy coats. Their thick necks, wedge-shaped heads and mobile snouts are used in feeding to uproot the ground and find prey or plant material. Feral pigs are easily distinguished from domestic pigs via a smaller leaner and more muscular stature, shorter hind quarters, longer snouts and tusks. Older boars usually develop a thick keratinous shield over their shoulders, which provides some protection during fights with other boars. Feral pig hair is longer and coarser than a domestic pigs and sometimes forms in a tuft along their back (hence, the name razorback). The tails of feral pigs are not curly as in domestic pigs, they are instead long and straight with a bushy tip.
Ecological characteristics of feral pig activity, group size and home range size should be considered in any management strategy aimed to control pig numbers or reduce their negative impact. Feral pig activity varies between different habitats and climates. High activity has been reported to occur in early morning and late afternoon in tropical climates (Diong 1982). However, in India pigs have been reported to feed nocturnally to raid croplands (Sekhar 1998, in Wolf and Conover 2003). On Santa Cruz Island (California) the milder weather of fall and late winter causes pigs to be more active in the morning and evening, while the short cool and often rainy days of winter causes midday activity. Pigs on the island were active at night mostly when conditions were warm and dry (Van Vuren 1984, in Wolf and Conover 2003).
In terms of group structure, in North-western Australia mob sizes are generally about 12 or less, although occasionally mobs of 30 pigs are seen. Adult boars are mostly solitary.. In South Carolina the average home range of boars is 226 hectares, while the average for sows is 181 hectares (Wood and Brenneman 1980, in Wolf and Conover 2003).Whereas in Austrailia average home range can vary from 140 hectares for a boar in Namagdi National park , Australian capital territory (McIlroy and Saillard 1989), to 430 hectares for a boar in Western New South Wale (Giles 1980).
Feral pigs are polyoestrous, adult sows have a 21 day oestrous cycle and a gestation period of 112-114 days (Choquenot et al.1996). Estimated litter size is 4.5-6.3 viable young per sow (Twigg et al. 2005, Choquenot et al. 1996) but in good conditions 10 piglets can be born to one sow.
Lifecycle Stages
Pigs are normally social animals but adult boars over 18 months old are invariably solitary (McIlroy 1990).
Uses
Captain Cook used the pig in trading with the natives as early as 1777. \"A small pig of 10 or 12 pounds\" was traded for a spike but a \"hog\" was exchanged for a hatchet (Cook 1784, in Diong 1982).\r\n
In central Europe the false spruce webworm (Cephalcia abietis) causes defoliation of Norway spruce trees; high densities of boars are able to cause high mortality to insect larvae by up to 70%, however they also cause damage to tree roots making the perceived benefit negligible (Fuhrer and Fischer 1991, in Wolf and Conover 2003).\r\n
In central Europe the false spruce webworm (Cephalcia abietis) causes defoliation of Norway spruce trees; high densities of boars are able to cause high mortality to insect larvae by up to 70%, however they also cause damage to tree roots making the perceived benefit negligible (Fuhrer and Fischer 1991, in Wolf and Conover 2003).\r\n
In many highland areas of New Guinea pigs are deliberately placed into gardens at the end of a harvest sequence and prior to gardening to remove remaining sweet potato tubers and to assist in turning and aerating the soil before replanting (Westermann 1968, Paglau 1982, Wood and Humphreys 1982, Tucker 1986, Kohun in hide 2003).
Habitat Description
The feral pig adapts to a variety of environments from Mediterranean oak woodland forests to the semi-arid rangelands of Eastern Australia, from the flood plains, billabongs and grassland savannas of tropical North-western Australia to the gray beech forests of the Smoky Mountains in America and from the wetland and lowland evergreen monsoon forests of Australia to the fresh water marshes and brackish water marshes of South Carolina (Wood and Brenneman 1980, in Wolf and Conover 2003). Wild pigs are rarely found over 1650m (Bulmer and Bulmer 1964, in hide 2003), but are known to be found at altitudes as high as 3000m in New Guinea (Flannery 1995, in Hide 2003).\r\n
Home ranges of pigs are smaller during the dry season than during the wet season. During the dry season on Santa Catalina pigs preferred cool moist canyon bottoms due to a physiological need for free water. Dense vegetation was more actively sought after than open areas such as grasslands (Baber and Coblentz 1986, in Wolf and Conover 2003). \r\n
The presence of crops in the near area (for example palm dates or oat hay cultivations) provide a food supplement and may greatly increase feral pig density; the close location of cereal crops in one study increased the density of feral pigs almost four-fold (Caley 1993, in Wolf and Conover 2003). Similarly the presence of adjacent palm cultivations in Malaysia was found to increase pigs density by 10 to 100 times (Ickes Paciorek and Thomas 2005). \r\n
High densities of pigs may also be attributed to water availability. The recent expansion in feral pig distribution in Australia has been attributed to the increase in suitable habitats, in particular, an increase in water availability from farm dams and developing forest industries (Spencer and Hampton 2005).
Reproduction
Feral pigs are polyoestrus: adult females have a 21-day oestrus cycle and a gestation period of about 112-114 days. In New Zealand they probably breed throughout the year, though mainly in spring and summer (Wodzicki 1950; J. McIlroy unpublished). Their litter size is usually between 6 and 10 piglets, but usually only half this number survives. They reach breeding age at between 10 and 12 months (Wodzicki 1950).
In one study females were found to have about 5 young every 0.86 years with some females having two litters per year. In this study fertility continued to increase with age until it peaked at two to three years of age. 58% of piglets died before weaning (Baber and Coblentz 1986, in Wolf and Conover 2003).
Nutrition
Pigs lack the multiple stomachs found in ruminants such as cattle and goats. Feral pigs are omnivores with an opportunistic diet, including high-fibre (> 25%) low-protein grasses, legumes, herbs and roots. They readily feed on crops, fallen fruits, seeds and small animals (McIlroy 1990). Pigs regularly root the ground in search of roots, fungus, nuts, seeds and grubs (Frederick 1998, Sicuro 2002, in Wolf and Conover 2003). In their native Mediterranean woodland the wild boar compensates for the reduced supply of acorns in the spring by raiding underground hoards of acorns collected and buried by small mammals (the availability of acorns is critical to female boars as they need the extra nutrition for lactation) (Focardi Capizzi and Monetti 2000, in Wolf and Conover 2003).
Pigs adapt their diet to best utilise local resources. In the semi-arid rangelands of eastern Australia and in New Guinea feral pigs will regularly hunt and devour lambs (particularly twin lambs (which are weaker) (Choquenot, Lukins and Curran 1997, in Wolf and Conover 2003; Hide 2003). On Horn Island, Mississippi, hogs take advantage of high seasonal abundances of insects, crabs and dead fish (Baron 1982, in Wolf and Conover 2003). On Santa Cruz Island, California, acorns and new growth of grasses and forbs are major components of the feral pig’s diet (Van Vuren 1984, in Wolf and Conover 2003).
In South Carolina fruits, especially acorns are the most common food type consumed in fall and winter; herbage and foliage are most common in the spring; roots are most common in the summer. Invertebrates and vertebrates are also consumed, though they were not as important. The consumption of woody plants may be underestimated in stomach contents surveys as the starches and sap obtained from the roots of such plants go undetected (Wood and Roark 1980, in Wolf and Conover 2003).
In the western South Texas Plains (introduced range) feral pigs have a spring-summer diet that consists mainly of vegetation, while acorns are their winter food source. Their autumn diet consists of roots and corn. Animal matter consisting of deer, morning doves, reptiles and other birds represents a small portion of the hog's diet. Of these, reptiles were the most susceptible to predation (Taylor and Hellgren 1997, in Wolf and Conover 2003).
In one study conducted in Hawaii by Diong 1982, food habits were characterised by (1) an omnivorous diet consisting mainly of plant matter, (2) a staple of tree ferns, (3) a seasonal switch from tree ferns to strawberry guava, and (4) a strong reliance of earthworms as a source of animal protein. The dietary range covered 40 plant species (63% herbaceous species, 33% trees and woody vine). Tree ferns were the most concentrated source of sugar and starch.
Pathway
Expansion into new areas can result from transport for hunting, escape from confined facilities, dispersal of wild populations and escape of domestic swine from free ranging commercial ranches (Gipson Hlavachick And Berger 1998, in Wolf and Conover 2003).Released as food.
Principal source:
Compiler: IUCN SSC Invasive Species Specialist Group
Updates with support from the Overseas Territories Environmental Programme (OTEP) project XOT603, a joint project with the Cayman Islands Government - Department of Environment
Review:
Publication date: 2010-05-18
Recommended citation: Global Invasive Species Database (2026) Species profile: Sus scrofa. Downloaded from http://www.iucngisd.org/gisd/speciesname/Sus+scrofa on 15-05-2026.
General Impacts
Please follow this link for details on the general impacts of S. scrofa compiled by the ISSG.
Management Info
Poisoning with sodium monofluoracetate (1080) is the most popular method used to control feral pigs. Most pigs vomit within four hours of ingestion. This may be potentially hazardous to nontarget organisms and may result in the survival of the pig. The use of anti-emetics such as metoclopramide, thiethylperazine and prochlorperazine may prevent vomiting at high doses (O'Brien et al. 1986, in Wolf and Conover 2003).\r\n
A vaccine for pseudorabies and swine brucellosis in fish meal bait may be used in late summer (when natural food supplies are low) to control these diseases (Fletcher et al. 1990, in Wolf and Conover 2003).\r\n
In the mid 1900s New Zealand conservation practitioners applied mainland hunting techniques to eradicate feral pig populations from small islands (<200 ha, Veitch and Bell, 1990, in Cruz et al. 2005). More recently poisoning techniques have been developed to control or eradicate feral pig populations (Choquenot et al., 1990; O'Brien and Lukins, 1990, in Cruz et al. 2005). Hunting and poisoning techniques used in combination, now facilitate pig eradication efforts on larger islands (Lombardo and Faulkner, 2000, Schuyler et al., 2002, Veitch and Bell, 1990, in Cruz et al. 2005).\r\n
In Hawaii, snaring has been used to control pigs within 600–800 km2 fenced enclosures located in remote areas of rain forest in the Haleakala National Park (Maui) (Anderson and Stone 1993). Many people place a high cultural value on pigs (ie: using them as a food convenient food source) so that removing them from designated areas may not be acceptable without a clear idea of the benefits. Snaring would is not always be an acceptable method of control. In addition, the fact that pigs are highly mobile means it is uneconomic for an individual landowners or controlling agency to control them (as pigs as they quickly move in from adjacent properties to replace the removed ones).\r\n
Much wisdom and insight can be gained from the case study of pig removal from Santiago Island in the Galapagos Archipelago (off the coast of Ecuador). Factors that proved critical to the successful eradication of the feral pig on the island were: (1) a sustained effort, (2) an effective poisoning campaign, (3) a hunting program, (4) access to animals by cutting more trails and, (5) an intensive monitoring program. Throughout the 1970s and 1980s, hunting effort was low (<500 hunter-days/year), while in the early 1990s effort increased but fluctuated. In contrast, the revised campaign in the mid-1990s resulted in a continuous, minimum annual effort of 1500 hunter-days/year. Hunter access to pigs was critical. Extra trails were cut and goats were not hunted in order to keep vegetation suppressed (allowing hunters and dogs access to all areas of the island). Motivating hunters was a continual challenge, especially when pigs were at low densities. However, social, moral boosting events and financial incentives maintained hunter motivation. While the poisoning campaign killed relatively few pigs compared to hunting, the low cost of the poisoning made such efforts especially cost-effective. The compounds used were toxic to most species, and thus the pros of using them for eradication had to be balanced with the potential impact on non-target species (Donlan et al., 2003a, in Cruz et al. 2005). In 2000, six months after the last pig was shot, the last pig was poisoned following an intensive monitoring effort. A sustained monitoring effort was critical to successful eradication. The lack of such an effort is responsible for many eradication failures (Campbell et al., 2004, in Cruz et al. 2005).
Countries (or multi-country features) with distribution records for Sus scrofa
ALIEN RANGE
- [1] american samoa
- [1] argentina
- [7] australia
- [1] bahamas
- [1] brazil
- [1] chile
- [7] cook islands
- [1] curacao
- [1] dominica
- [1] dominican republic
- [2] ecuador
- [4] fiji
- [1] france
- [8] french polynesia
- [1] french southern territories
- [1] guam
- [1] india
- [1] jamaica
- [9] kiribati
- [1] marshall islands
- [2] mauritius
- [1] mayotte
- [1] mexico
- [3] micronesia, federated states of
- [1] montserrat
- [1] nauru
- [7] new caledonia
- [1] new guinea
- [8] new zealand
- [1] niue
- [4] northern mariana islands
- [1] pakistan
- [1] palau
- [13] papua new guinea
- [1] pitcairn
- [1] puerto rico
- [1] reunion
- [1] saint lucia
- [2] samoa
- [8] solomon islands
- [1] south america
- [1] tonga
- [22] united states
- [1] virgin islands, u.s.
- [1] wallis and futuna
NATIVE RANGE
- malaysia
- spain
Informations on Sus scrofa has been recorded for the following locations. Click on the name for additional informations.
Lorem Ipsum
| Location | Status | Invasiveness | Occurrence | Source |
Details of Sus scrofa in information
Status
Invasiveness
Arrival date
Occurrence
Source
Introduction
Species notes for this location
Location note
Management notes for this location
Method:
Source:
Start date:
End date:
Impact
 Mechanism:
Mechanism:  Outcome:
Outcome:  Ecosystem services:
Ecosystem services: Impact information
Please follow this link for details on the general impacts of S. scrofa compiled by the ISSG.
Red List assessed species 281: EX = 7; EW = 5; CR = 109; EN = 81; VU = 54; LR/nt = 1; NT = 14; DD = 1; LC = 9;
- Abutilon sandwicense CR
- Acacia koaia VU
- Alectryon macrococcus CR
- Alphitonia ponderosa VU
- Alsinidendron lychnoides CR
- Alsinidendron obovatum CR
- Alsinidendron trinerve CR
- Alsinidendron viscosum CR
- Anas aucklandica VU
- Anas wyvilliana EN
- Aphelocoma insularis NT
- Apteryx haastii VU
- Araucaria hunsteinii LR/nt
- Argyroxiphium kauense CR
- Argyroxiphium sandwicense VU
- Astelia waialealae CR
- Bidens conjuncta VU
- Bidens cosmoides EN
- Bidens populifolia VU
- Bobea sandwicensis VU
- Bonamia menziesii CR
- Branta sandvicensis VU
- Bulimulus darwini VU
- Buteo solitarius NT
- Calamagrostis expansa VU
- Calamagrostis hillebrandii EN
- Callerya neocaledonica CR
- Camarhynchus pauper CR
- Canavalia molokaiensis CR
- Caretta caretta EN
- Casuarius bennetti NT
- Cenchrus agrimonioides CR
- Chamaesyce deppeana CR
- Chamaesyce halemanui CR
- Chamaesyce remyi CR
- Chamaesyce rockii CR
- Chamaesyce sparsiflora VU
- Charpentiera densiflora CR
- Cheirodendron dominii EN
- Chelonia mydas EN
- Christella boydiae EN
- Clermontia calophylla EN
- Clermontia drepanomorpha EN
- Clermontia hawaiiensis VU
- Clermontia lindseyana EN
- Clermontia peleana EW
- Clermontia pyrularia CR
- Clermontia tuberculata EN
- Clermontia waimeae EN
- Coccyzus ferrugineus VU
- Coenocorypha aucklandica NT
- Colubrina oppositifolia CR
- Ctenitis squamigera CR
- Cyanea acuminata CR
- Cyanea asarifolia CR
- Cyanea asplenifolia CR
- Cyanea crispa CR
- Cyanea dunbariae CR
- Cyanea eleeleensis CR
- Cyanea glabra CR
- Cyanea horrida CR
- Cyanea pinnatifida EW
- Cyanea st-johnii CR
- Cyanea superba EW
- Cyanea truncata EW
- Cyclura collei CR
- Cyclura cornuta VU
- Cyclura stejnegeri EN
- Cyrtandra giffardii EN
- Cyrtandra kaulantha CR
- Cyrtandra polyantha CR
- Cyrtandra waiolani EW
- Dasyornis brachypterus EN
- Dermochelys coriacea CR
- Diomedea antipodensis VU
- Diomedea dabbenena CR
- Diomedea epomophora VU
- Diploglossus montisserrati CR
- Ducula galeata EN
- Emoia adspersa EN
- Engaeus martigener EN
- Engaeus urostrictus VU
- Engaewa similis LC
- Engaewa walpolea EN
- Epicrates monensis EN
- Eretmochelys imbricata CR
- Erythrura gouldiae EN
- Euastacus armatus DD
- Euastacus australasiensis LC
- Euastacus balanesis EN
- Euastacus bidawalis EN
- Euastacus bindal CR
- Euastacus brachythorax EN
- Euastacus clarkae CR
- Euastacus claytoni EN
- Euastacus crassus EN
- Euastacus dalagarbe CR
- Euastacus dharawalus CR
- Euastacus diversus EN
- Euastacus eungella CR
- Euastacus fleckeri EN
- Euastacus gamilaroi CR
- Euastacus girurmulayn CR
- Euastacus gumar EN
- Euastacus guruhgi CR
- Euastacus guwinus CR
- Euastacus hirsutus EN
- Euastacus hystricosus EN
- Euastacus jagabar CR
- Euastacus jagara CR
- Euastacus maccai EN
- Euastacus maidae CR
- Euastacus mirangudjin CR
- Euastacus monteithorum CR
- Euastacus pilosus EN
- Euastacus polysetosus EN
- Euastacus rieki EN
- Euastacus robertsi CR
- Euastacus setosus CR
- Euastacus simplex VU
- Euastacus spinichelatus EN
- Euastacus sulcatus VU
- Euastacus suttoni VU
- Euastacus urospinosus EN
- Euastacus valentulus LC
- Euastacus wiowuru NT
- Euastacus yanga LC
- Euastacus yarreansis VU
- Euastacus yigara CR
- Eugenia koolauensis EN
- Euphorbia haeleeleana EN
- Gallicolumba salamonis EX
- Gallicolumba sanctaecrucis EN
- Gallinula nesiotis VU
- Gallinula pacifica CR
- Gallirallus lafresnayanus CR
- Gallirallus sylvestris EN
- Gardenia mannii CR
- Geocrinia vitellina VU
- Gouania vitifolia CR
- Gymnomyza aubryana CR
- Hemignathus lucidus CR
- Hemignathus parvus VU
- Hesperomannia arborescens CR
- Hesperomannia arbuscula CR
- Hibiscadelphus woodii CR
- Hibiscus clayi CR
- Himantoglossum adriaticum LC
- Hypericum corsicum LC
- Icterus oberi CR
- Labordia cyrtandrae CR
- Laterallus spilonotus VU
- Leptodactylus fallax CR
- Lewinia muelleri VU
- Lioscincus steindachneri EN
- Litoria dayi EN
- Litoria lorica CR
- Litoria nannotis EN
- Litoria nyakalensis CR
- Litoria pearsoniana NT
- Litoria rheocola EN
- Lonchura stygia NT
- Loxops coccineus EN
- Marmorosphax boulinda VU
- Marmorosphax kaala CR
- Marmorosphax montana VU
- Marmorosphax taom CR
- Marmorosphax tricolor LC
- Mastacomys fuscus NT
- Masticophis anthonyi CR
- Megacrex inepta NT
- Megalurus albolimbatus VU
- Megapodius laperouse EN
- Megapodius pritchardii EN
- Melamprosops phaeosoma CR
- Melicope balloui EN
- Melicope saint-johnii EN
- Mergus australis EX
- Metrosideros bartlettii EN
- Mimus macdonaldi VU
- Mimus trifasciatus CR
- Mixophyes fleayi EN
- Moho bishopi EX
- Moho braccatus EX
- Myadestes lanaiensis CR
- Myadestes myadestinus EX
- Myadestes obscurus VU
- Myadestes palmeri CR
- Nannoscincus garrulus EN
- Nannoscincus hanchisteus CR
- Nannoscincus manautei CR
- Nannoscincus rankini VU
- Nesotriccus ridgwayi VU
- Nothocestrum peltatum CR
- Numenius tahitiensis VU
- Oedodera marmorata CR
- Oreomystis bairdi CR
- Oreomystis mana EN
- Palmeria dolei CR
- Paroreomyza montana EN
- Pelagodoxa henryana CR
- Phaeognathus hubrichti EN
- Phalacrocorax colensoi VU
- Phalacrocorax featherstoni EN
- Phalacrocorax onslowi CR
- Phalanger alexandrae EN
- Phlegmariurus nutans CR
- Phylloscopus amoenus VU
- Phyllostegia kaalaensis CR
- Phyllostegia mollis CR
- Pinaroloxias inornata VU
- Potamon fluviatile NT
- Potorous longipes EN
- Pritchardia affinis CR
- Pritchardia glabrata EN
- Pritchardia kaalae CR
- Pritchardia lanaiensis EN
- Pritchardia lanigera EN
- Pritchardia limahuliensis CR
- Pritchardia napaliensis CR
- Pritchardia perlmanii EN
- Pritchardia viscosa CR
- Procellaria conspicillata VU
- Procellaria parkinsoni VU
- Psephotus chrysopterygius EN
- Pseudobulweria rostrata NT
- Pseudonestor xanthophrys CR
- Pseudophryne pengilleyi EN
- Psittacula eques EN
- Psittirostra psittacea CR
- Pteralyxia kauaiensis EN
- Pterodroma arminjoniana VU
- Pterodroma axillaris EN
- Pterodroma brevipes VU
- Pterodroma caribbaea CR
- Pterodroma hasitata EN
- Pterodroma leucoptera VU
- Pterodroma magentae CR
- Pterodroma phaeopygia CR
- Pterodroma sandwichensis VU
- Pterodroma solandri VU
- Pteropus mariannus EN
- Puffinus auricularis CR
- Puffinus bulleri VU
- Puffinus huttoni EN
- Puffinus newelli EN
- Rhacodactylus auriculatus LC
- Rheobatrachus silus EX
- Rhionaeschna galapagoensis EN
- Rhynchomeles prattorum EN
- Rhynochetos jubatus EN
- Schiedea kaalae CR
- Setonix brachyurus VU
- Simiscincus aurantiacus VU
- Sus cebifrons CR
- Sus oliveri EN
- Sus philippensis VU
- Sylvilagus graysoni EN
- Tacheocampylaea cyrniaca EN
- Tacheocampylaea raspailii VU
- Tacheocampylaea romagnolii CR
- Taudactylus acutirostris CR
- Taudactylus diurnus EX
- Taudactylus pleione CR
- Taudactylus rheophilus CR
- Tetraplasandra gymnocarpa CR
- Thalassarche steadi NT
- Thylogale calabyi EN
- Tinostoma smaragditis EN
- Todiramphus farquhari NT
- Todiramphus godeffroyi CR
- Todiramphus ruficollaris VU
- Trigonostemon cherrieri CR
- Turnix melanogaster VU
- Typhlops biminiensis NT
- Urera kaalae CR
- Urosaurus clarionensis VU
- Vestiaria coccinea VU
- Vini ultramarina EN
- Xantusia riversiana LC
- Xylosma crenatum CR
Locations

 AUSTRALIAFRENCH POLYNESIAMONTSERRATNEW CALEDONIAUNITED STATES
AUSTRALIAFRENCH POLYNESIAMONTSERRATNEW CALEDONIAUNITED STATESMechanism
[1] Competition
Competition- NEW CALEDONIA
[3] Predation
Predation- FRENCH POLYNESIA
- MONTSERRAT
- NEW CALEDONIA
[4] Grazing/Herbivory/Browsing
Grazing/Herbivory/Browsing- FRENCH POLYNESIA
- MONTSERRAT
- NEW CALEDONIA
[3] Rooting/Digging
Rooting/Digging- FRENCH POLYNESIA
- MONTSERRAT
- UNITED STATES
[2] Trampling
Trampling- FRENCH POLYNESIA
- MONTSERRAT
[1] Interaction with other invasive species
Interaction with other invasive species- FRENCH POLYNESIA
Outcomes
[9] Environmental Ecosystem - Habitat
Environmental Ecosystem - Habitat- [1] Modification of hydrology/water regulation, purification and quality /soil moisture
- [5] Reduction in native biodiversity
- [2] Habitat degradation
- [1] Modification of successional patterns
[1] Socio-Economic
Socio-Economic- [1] Damage to forestry
Management information
Poisoning with sodium monofluoracetate (1080) is the most popular method used to control feral pigs. Most pigs vomit within four hours of ingestion. This may be potentially hazardous to nontarget organisms and may result in the survival of the pig. The use of anti-emetics such as metoclopramide, thiethylperazine and prochlorperazine may prevent vomiting at high doses (O'Brien et al. 1986, in Wolf and Conover 2003).\r\n
A vaccine for pseudorabies and swine brucellosis in fish meal bait may be used in late summer (when natural food supplies are low) to control these diseases (Fletcher et al. 1990, in Wolf and Conover 2003).\r\n
In the mid 1900s New Zealand conservation practitioners applied mainland hunting techniques to eradicate feral pig populations from small islands (<200 ha, Veitch and Bell, 1990, in Cruz et al. 2005). More recently poisoning techniques have been developed to control or eradicate feral pig populations (Choquenot et al., 1990; O'Brien and Lukins, 1990, in Cruz et al. 2005). Hunting and poisoning techniques used in combination, now facilitate pig eradication efforts on larger islands (Lombardo and Faulkner, 2000, Schuyler et al., 2002, Veitch and Bell, 1990, in Cruz et al. 2005).\r\n
In Hawaii, snaring has been used to control pigs within 600–800 km2 fenced enclosures located in remote areas of rain forest in the Haleakala National Park (Maui) (Anderson and Stone 1993). Many people place a high cultural value on pigs (ie: using them as a food convenient food source) so that removing them from designated areas may not be acceptable without a clear idea of the benefits. Snaring would is not always be an acceptable method of control. In addition, the fact that pigs are highly mobile means it is uneconomic for an individual landowners or controlling agency to control them (as pigs as they quickly move in from adjacent properties to replace the removed ones).\r\n
Much wisdom and insight can be gained from the case study of pig removal from Santiago Island in the Galapagos Archipelago (off the coast of Ecuador). Factors that proved critical to the successful eradication of the feral pig on the island were: (1) a sustained effort, (2) an effective poisoning campaign, (3) a hunting program, (4) access to animals by cutting more trails and, (5) an intensive monitoring program. Throughout the 1970s and 1980s, hunting effort was low (<500 hunter-days/year), while in the early 1990s effort increased but fluctuated. In contrast, the revised campaign in the mid-1990s resulted in a continuous, minimum annual effort of 1500 hunter-days/year. Hunter access to pigs was critical. Extra trails were cut and goats were not hunted in order to keep vegetation suppressed (allowing hunters and dogs access to all areas of the island). Motivating hunters was a continual challenge, especially when pigs were at low densities. However, social, moral boosting events and financial incentives maintained hunter motivation. While the poisoning campaign killed relatively few pigs compared to hunting, the low cost of the poisoning made such efforts especially cost-effective. The compounds used were toxic to most species, and thus the pros of using them for eradication had to be balanced with the potential impact on non-target species (Donlan et al., 2003a, in Cruz et al. 2005). In 2000, six months after the last pig was shot, the last pig was poisoned following an intensive monitoring effort. A sustained monitoring effort was critical to successful eradication. The lack of such an effort is responsible for many eradication failures (Campbell et al., 2004, in Cruz et al. 2005).
Locations
AUSTRALIACHILEECUADORFRANCEMAURITIUSMEXICOMONTSERRATNEW CALEDONIANEW ZEALANDNORTHERN MARIANA ISLANDSUNITED STATESManagement Category
Eradication- AUSTRALIA
- FRANCE
- NEW ZEALAND
- NORTHERN MARIANA ISLANDS
- UNITED STATES
Control- AUSTRALIA
- CHILE
- ECUADOR
- MAURITIUS
- MONTSERRAT
- NEW CALEDONIA
- NEW ZEALAND
- UNITED STATES
None- MEXICO
Unknown- FRANCE
- NEW ZEALAND
- UNITED STATES
Bibliography
90 references found for Sus scrofa
Management information
Management information
Anderson, S. J., Stone, C. P. 1993. Snaring to control feral pigs Sus scrofa in a remote Hawaiian rain forest. Biological Conservation 63: 195�201.
Atkinson, I. A. E. and Atkinson, T. J. 2000. Land vertebrates as invasive species on islands served by the South Pacific Regional Environment Programme. In: Invasive Species in the Pacific: A Technical Review and Draft Regional Strategy. South Pacific Regional Environment Programme, Samoa: 19-84.
Summary: This report reviews available information on the adverse effects of 14 alien vertebrates considered to be �significant invasive species on islands of the South Pacific and Hawaii, supplementing the authors� experience with that of other workers.
Summary: This report reviews available information on the adverse effects of 14 alien vertebrates considered to be �significant invasive species on islands of the South Pacific and Hawaii, supplementing the authors� experience with that of other workers.
Bomford, M., 2003. Risk Assessment for the Import and Keeping of Exotic Vertebrates in Australia. Bureau of Rural Sciences, Canberra.
Summary: Available from: http://www.feral.org.au/wp-content/uploads/2010/03/PC12803.pdf [Accessed August 19 2010]
Summary: Available from: http://www.feral.org.au/wp-content/uploads/2010/03/PC12803.pdf [Accessed August 19 2010]
Brown, Christopher E.; Pezeshki, S. Reza, 2000. A study on waterlogging as a potential tool to control Ligustrum sinense populations in western Tennessee. Wetlands. 20(3). September, 2000. 429-437.
Center for Aquatic and Invasive Plants, University of Florida (IFAS), 2010. Chinese privet: Ligustrum sinense
Summary: Available from: http://plants.ifas.ufl.edu/node/231 [Accessed 10 March 2010]
Summary: Available from: http://plants.ifas.ufl.edu/node/231 [Accessed 10 March 2010]
Cruz, F., Donlan, C.J., Campbell, K. and Carrion, V. 2005. Conservation action in the Galàpagos: feral pig (Sus scrofa) eradication from Santiago Island, Biological Conservation 121(3). [Accessed February 23 2006, from ScienceDirect (online database)]
Cuthbert, R. 2002. The role of introduced mammals and inverse density-dependent predation in the conservation of Hutton s shearwater, Biological Conservation 108(1). [Accessed 23 February 2006, from ScienceDirect (online database)]
Department of Primary Industries, Victoria, 2009. Impact Assessment - Small-leaf Privet (Ligustrum sinense) in Victoria
Summary: Available from: http://www.dse.vic.gov.au/dpi/vro/vrosite.nsf/pages/impact_small_leaved_privet [Accessed 10 March 2010]
Summary: Available from: http://www.dse.vic.gov.au/dpi/vro/vrosite.nsf/pages/impact_small_leaved_privet [Accessed 10 March 2010]
Department of Primary Industries, Victoria, 2009. Invasiveness Assessment - Small-leaf Privet (Ligustrum sinense) in Victoria
Summary: Available from: http://dpi.vic.gov.au/dpi/vro/vrosite.nsf/pages/invasive_small_leaf_privet [Accessed 10 March 2010]
Summary: Available from: http://dpi.vic.gov.au/dpi/vro/vrosite.nsf/pages/invasive_small_leaf_privet [Accessed 10 March 2010]
Dexter, N. 2003. Stochastic models of foot and mouth disease in feral pigs in the Australian semi-arid rangelandsJournal of Applied Ecology 40(2).
Ding, Jianqing; Reardon, Richard; Wu, Yun; Zheng, Hao; Fu, Weidong, 2006. Biological control of invasive plants through collaboration between China and the United States of America: a perspective. Biological Invasions. 8(7). OCT 2006. 1439-1450
Early Detection and Distribution Mapping System (EDDMapS), 2010. Chinese privet Ligustrum sinense Lour.
Summary: Available from: http://www.eddmaps.org/distribution/state.cfm?sub=3035&id=us_tx [Accessed 10 March 2010]
Summary: Available from: http://www.eddmaps.org/distribution/state.cfm?sub=3035&id=us_tx [Accessed 10 March 2010]
Engeman, R.M., Smith, H.T., Severson, R., Severson, M.A., Shwiff, A., Constantin, B. and Griffin, D. 2004. The Amount and Economic Cost of Feral Swine Damage to the Last Remnant of a Basin Marsh System in Florida, Journal for Nature Conservation 12: 143-147. [Accessed 1 March 2006, from ScienceDirect (online database)]
Engeman, R.M., Smith, H.T., Severson, R., Severson, M.A., Woodland, J., Shwiff, A., Constantin, B. and Griffin, D. 2004. Damage Reduction Estimates and Benefit-cost Ratios for Feral Swine Control From the Last Remnant of a Basin Marsh System in Florida, Environmental Conservation 31 (3): 207-211.
Greene, Brian T. and Bernd Blossey, August 5, 2009. COS 58-1: Patterns of privet: Why is the invasive plant Ligustrum sinense Lour associated with urban watersheds in the southeastern United States. Wednesday, August 5, 2009 - 8:00 AM. 94th ESA Annual Meeting Sunday August 2 -Friday, August 7 2009 Albuquerque, New Mexico.
Summary: Available from: http://eco.confex.com/eco/2009/techprogram/P18256.HTM [Accessed 10 March 2010]
Summary: Available from: http://eco.confex.com/eco/2009/techprogram/P18256.HTM [Accessed 10 March 2010]
Hampton, J.O., Spencer, P.B.S., Alpers, D.L., Twigg, L.E, Woolnough, A.P., Doust, J., Higgs, T. and Pluske, J. 2004. Molecular techniques, wildlife management and the importance of genetic population structure and dispersal: a case study with feral pigs, Journal of Applied Ecology 41.
Hanula, James L.; Scott Horn, and John W. Taylor, 2009. Chinese Privet (Ligustrum sinense) Removal and its Effect on Native Plant Communities of Riparian Forests. Invasive Plant Science and Management 2009 2:292-300
Summary: Available from: http://www.srs.fs.usda.gov/pubs/ja/ja_hanula018.pdf [Accessed 10 March 2010]
Summary: Available from: http://www.srs.fs.usda.gov/pubs/ja/ja_hanula018.pdf [Accessed 10 March 2010]
Harrington, Timothy B.; Miller, James H., 2005. Effects of application rate, timing, and formulation of glyphosate and triclopyr on control of Chinese privet (Ligustrum sinense). Weed Technology. 19(1). JAN-MAR 2005. 47-54.
Hide, R. 2003. Pig husbandry in New Guinea. A literature review and bibliography. Australian Centre for International Agricultural Research.
Ickes, K., Paciorek, C.J. and Thomas, S.C. 2005. Impacts of Nest Construction by Native Pigs (Sus scrofa) on Lowland Malaysian Rain Forest Saplings, Ecology 86(6). [Accessed 22 February 2006, from Ecological Society of America (online database)]
IUCN 2010. IUCN Red List of Threatened Species. Version 2010.4.
Summary: The IUCN Red List of Threatened Species provides taxonomic, conservation status and distribution information on taxa that have been globally evaluated using the IUCN Red List Categories and Criteria. This system is designed to determine the relative risk of extinction, and the main purpose of the IUCN Red List is to catalogue and highlight those taxa that are facing a higher risk of global extinction (i.e. those listed as Critically Endangered, Endangered and Vulnerable). The IUCN Red List also includes information on taxa that are categorized as Extinct or Extinct in the Wild; on taxa that cannot be evaluated because of insufficient information (i.e. are Data Deficient); and on taxa that are either close to meeting the threatened thresholds or that would be threatened were it not for an ongoing taxon-specific conservation programme (i.e. are Near Threatened).
Available from: http://www.iucnredlist.org/ [Accessed 25 May 2011]
Summary: The IUCN Red List of Threatened Species provides taxonomic, conservation status and distribution information on taxa that have been globally evaluated using the IUCN Red List Categories and Criteria. This system is designed to determine the relative risk of extinction, and the main purpose of the IUCN Red List is to catalogue and highlight those taxa that are facing a higher risk of global extinction (i.e. those listed as Critically Endangered, Endangered and Vulnerable). The IUCN Red List also includes information on taxa that are categorized as Extinct or Extinct in the Wild; on taxa that cannot be evaluated because of insufficient information (i.e. are Data Deficient); and on taxa that are either close to meeting the threatened thresholds or that would be threatened were it not for an ongoing taxon-specific conservation programme (i.e. are Near Threatened).
Available from: http://www.iucnredlist.org/ [Accessed 25 May 2011]
IUCN/SSC Invasive Species Specialist Group (ISSG)., 2010. A Compilation of Information Sources for Conservation Managers.
Summary: This compilation of information sources can be sorted on keywords for example: Baits & Lures, Non Target Species, Eradication, Monitoring, Risk Assessment, Weeds, Herbicides etc. This compilation is at present in Excel format, this will be web-enabled as a searchable database shortly. This version of the database has been developed by the IUCN SSC ISSG as part of an Overseas Territories Environmental Programme funded project XOT603 in partnership with the Cayman Islands Government - Department of Environment. The compilation is a work under progress, the ISSG will manage, maintain and enhance the database with current and newly published information, reports, journal articles etc.
Summary: This compilation of information sources can be sorted on keywords for example: Baits & Lures, Non Target Species, Eradication, Monitoring, Risk Assessment, Weeds, Herbicides etc. This compilation is at present in Excel format, this will be web-enabled as a searchable database shortly. This version of the database has been developed by the IUCN SSC ISSG as part of an Overseas Territories Environmental Programme funded project XOT603 in partnership with the Cayman Islands Government - Department of Environment. The compilation is a work under progress, the ISSG will manage, maintain and enhance the database with current and newly published information, reports, journal articles etc.
Kessler, C.C., 2002. Eradication of feral goats and pigs and consequences for other biota on Sarigan Island, Commonwealth of the Northern Mariana Islands. In Turning the tide: the eradication of invasive species: 132-140. Veitch, C.R. and Clout, M.N.(eds). IUCN SSC Invasive Species Specialist Group. IUCN. Gland. Switzerland and Cambridge. UK.
Summary: Eradication case study in Turning the tide: the eradication of invasive species.
Summary: Eradication case study in Turning the tide: the eradication of invasive species.
Marvin, David C.; Bradley, Bethany A.; Wilcove, David S., 2009. A Novel, Web-based, Ecosystem Mapping Tool Using Expert Opinion. Natural Areas Journal. 29(3). JUL 2009. 281-292.
Matlack, Glenn R., 2002. Exotic plant species in Mississippi, USA: Critical issues in management and research. Natural Areas Journal. 22(3). July, 2002. 241-247.
Mauremootoo, J. R.; C. G. Jones, W. A. Strahm, M. E. Dulloo, and Y. Mungroo., 2002. The effectiveness of weeded and fenced Conservation Management Areas as a means of maintaining the threateneed biodiversity of mainland Mauritius. In Turning the tide: the eradication of invasive species: 406 - 414. IUCN SSC Invasive Species Specialist Group. IUCN. Gland. Switzerland and Cambridge. UK.
Summary: Eradication case study In Turning the tide: the eradication of invasive species.
Summary: Eradication case study In Turning the tide: the eradication of invasive species.
Montaldo, Norberto H., 1993. Avian dispersal and reproductive success of two species of Ligustrum (Oleaceae) in a subtropical forest relict in Argentina. Revista Chilena de Historia Natural. 66(1). 1993. 75-85.
Moors, P. J., Atkinson, I. A. E. and Sherley, G. H. 1992. Reducing the rat threat to island birds. Bird Conservation International 2: 93�114.
Nel, J.L.; D.M. Richardson; M. Rouget; T.N. Mgidi; N. Mdzeke; D.C. Le Maitre; B.W. van Wilgen; L. Schonegevel; L. Henderson and S. Neser, 2004. A proposed classification of invasive alien plant species in South Africa: towards prioritizing species and areas for management action. Working for Water South African Journal of Science 100, January/February 2004
Summary: Available from: http://www.dwaf.gov.za/WFW/Docs/Papers/SAJSFeb2004nel.pdf [Accessed 10 March 2010]
Summary: Available from: http://www.dwaf.gov.za/WFW/Docs/Papers/SAJSFeb2004nel.pdf [Accessed 10 March 2010]
Pacific Island Ecosystems at Risk (PIER), 2005. Risk Assessment: Ligustrum sinense Lour., Oleaceae
Summary: Available from: http://www.hear.org/pier/wra/pacific/ligustrum_sinense_htmlwra.htm [Accessed 10 March 2010]
Summary: Available from: http://www.hear.org/pier/wra/pacific/ligustrum_sinense_htmlwra.htm [Accessed 10 March 2010]
Pacific Island Ecosystems at Risk (PIER), 2010. Ligustrum sinense Lour., Oleaceae
Summary: Available from: http://www.hear.org/pier/species/ligustrum_sinense.htm [Accessed 10 March 2010]
Summary: Available from: http://www.hear.org/pier/species/ligustrum_sinense.htm [Accessed 10 March 2010]
Patry, M. 2001. Feral Pig Eradication Campaign on Santiago Island, Galapagos. Informe Galapagos 2001. Fundaci�n Natura, Quito (in press).
Summary: Describes the eradication project for pigs on Santiago Island, Galapagos. Covers the entire project from pre-1997 to May 2001.
Summary: Describes the eradication project for pigs on Santiago Island, Galapagos. Covers the entire project from pre-1997 to May 2001.
Queensland Pest Animal Strategy, 2004. Feral Pigs, The State of Queensland (Natural Resouces, Mines and Energy).
Summary: The feral pig management strategy outlines the best practises for the management of feral pigs to minimise their impact on the environment, economy and health of Queensland.
Summary: The feral pig management strategy outlines the best practises for the management of feral pigs to minimise their impact on the environment, economy and health of Queensland.
Schuyler, P. T., Garcelon, D. K. and Escover, S., 2002. Eradication of feral pigs (Sus scrofa) on Santa Catalina Island, California, USA. In Turning the tide: the eradication of invasive species: 274-286. Veitch, C.R. and Clout, M.N.(eds). IUCN SSC Invasive Species Specialist Group. IUCN. Gland. Switzerland and Cambridge. UK.
Summary: Eradication case study in Turning the tide: the eradication of invasive species.
Summary: Eradication case study in Turning the tide: the eradication of invasive species.
Smith, K. E.; G. B Runion; S.A. Prior; A. J. Price; H. H. Rogers and H. A. Torbert, 2008. Chinese Privet (Ligustrum sinense) in an elevated CO2 Environment. Botany Research Journal 1 (2): 43-48, 2008.
Summary: Available from: http://www.medwellonline.net/fulltext/brj/2008/43-48.pdf [Accessed 10 March 2010]
Summary: Available from: http://www.medwellonline.net/fulltext/brj/2008/43-48.pdf [Accessed 10 March 2010]
Spencer, P.B.S. and Hampton, J.O. 2005. Illegal Translocation and Genetic Structure of Feral Pigs in Western Australia, Journal of Wildlife Management 69(1). [Accessed 23 February 2006, from BioOne (online database)]
Thetford, Mack; Berry, James B., 2000. Response of five woody landscape plants to Primo and pruning. Journal of Environmental Horticulture. 18(3). September, 2000. 132-136.
Twigg, L.E., Lowe, T., Martin, G. and Michael, E. 2005. Feral Pigs in North-western Australia: Basic Biology, Bait Consumption, and the Efficacy of 1080 Baits, Wildlife Research 32: 281-296.
Ulyshen, Michael D.; Scott Horn and James L. Hanula, 2009. Response of beetles (Coleoptera) at three heights to the experimental removal of an invasive shrub, Chinese privet (Ligustrum sinense), from floodplain forests. Biological Invasions DOI 10.1007/s10530-009-9569-2
Varnham, K. 2006. Non-native species in UK Overseas Territories: a review. JNCC Report 372. Peterborough: United Kingdom.
Summary: This database compiles information on alien species from British Overseas Territories.
Available from: http://www.jncc.gov.uk/page-3660 [Accessed 10 November 2009]
Summary: This database compiles information on alien species from British Overseas Territories.
Available from: http://www.jncc.gov.uk/page-3660 [Accessed 10 November 2009]
Vidra, Rebecca L.; Shear, Theodore H.; Stucky, Jon M., 2007. Effects of vegetation removal on native understory recovery in an exotic-rich urban forest. Journal of the Torrey Botanical Society. 134(3). JUL-SEP 2007. 410-419.
West., C. J., 2002. Eradication of alien plants on Raoul Island, Kermadec Islands, New Zealand. In Turning the tide: the eradication of invasive species: 381-388. Veitch, C.R. and Clout, M.N.(eds). IUCN SSC Invasive Species Specialist Group. IUCN. Gland.
Summary: Eradication case study in Turning the tide: the eradication of invasive species.
Summary: Eradication case study in Turning the tide: the eradication of invasive species.
Williams, Peter A.; Karl, Brian J.; Bannister, Peter; Lee, William G., 2000. Small mammals as potential seed dispersers in New Zealand. Austral Ecology. 25(5). October, 2000. 523-532.
Williams, Rick & Patrick Minogue, 2008. Biology and Management of Chinese Privet. FR189, one of a series of the School of Forest Resources and Conservation Department, Florida Cooperative Extension Service, Institute of Food and Agricultural Sciences, University of Florida
Summary: Available from: http://edis.ifas.ufl.edu/pdffiles/FR/FR25000.pdf [Accessed 10 March 2010]
Summary: Available from: http://edis.ifas.ufl.edu/pdffiles/FR/FR25000.pdf [Accessed 10 March 2010]
Wirth, Ferdinand F.; Davis, Kathy J.; Wilson, Sandra B., 2004. Florida nursery sales and economic impacts of 14 potentially invasive landscape plant species. Journal of Environmental Horticulture. 22(1). March 2004. 12-16
Wodzicki, K. A. 1950. Introduced mammals of New Zealand. Department of Scientific and Industrial Research Bulletin 98. Department of Scientific and Industrial Research, Wellington.
Wolf, T. and Conover, M.R. 2003. Feral Pigs and the Environment: An Annotated Bibliography. Jack H. Berryman Institute 21: Utah State University, Logan; Mississippi State University, Starkville.
Zhang, Yan-Zhuo; Hanula, James L.; Sun, Jiang-Hua, 2008. Survey for potential insect biological control agents of Ligustrum sinense (Scrophulariales : Oleaceae) in China. Florida Entomologist. 91(3). SEP 2008. 372-382.
Zhang, Yanzhuo; Sun, Jianghua; Hanula, James L., 2009. Biology and Life History of Argopistes tsekooni (Coleoptera: Chrysomelidae) in China, a Promising Biological Control Agent of Chinese Privet. Annals of the Entomological Society of America. 102(3). MAY 2009. 508-516
General information
Aplet, G. H., S. J. Anderson, and C. P. Stone. 1991. Association between feral pig disturbance and the composition of some alien plant assemblages in Hawaii Volcanos National Park. Vegetation 95:55-62.
Auld, B. A. and Tisdell, C. A. 1986. In Groves, R. H. and Burdon, J. J. (eds.) Ecology of Biological Invasions: An Australian Perspective. Australian Academy of Science, Canberra.
Barthelat, pers. comm., 2007
Summary: Personal communication with Fabien Barthelat, an expert of flora of Mayotte.
Summary: Personal communication with Fabien Barthelat, an expert of flora of Mayotte.
Brescia, F. 2005. Am�lioration des connaissances sur l��cologie des bulimes (dynamique des populations, pr�dation), �tude des pr�l�vements dans les stocks naturels et poursuite du transfert de la m�thode d��levage sur l��le des Pins. Rapport d��tude IAC-DRN (Province Sud). Programme Elevage et Faune Sauvage IAC.
Burton, Michele L.; Samuelson, Lisa J., 2008. Influence of urbanization on riparian forest diversity and structure in the Georgia Piedmont, US. Plant Ecology. 195(1). MAR 2008. 99-115.
Burton, Michele L.; Samuelson, Lisa J.; Pan, Shufen, 2005. Riparian woody plant diversity and forest structure along an urban-rural gradient. Urban Ecosystems. 8(1). MAR 2005. 93-106.
Butaud and Meyer, pers. comm. 2007
Summary: Personal communication with Jean Fran�ois Butaud and Jean Yves Meyer, two experts on flora and fauna of French Polynesia
Summary: Personal communication with Jean Fran�ois Butaud and Jean Yves Meyer, two experts on flora and fauna of French Polynesia
CONABIO. 2008. Sistema de informaci�n sobre especies invasoras en M�xico. Especies invasoras - Mam�feros. Comisi�n Nacional para el Conocimiento y Uso de la Biodiversidad. Fecha de acceso.
Summary: English:
The species list sheet for the Mexican information system on invasive species currently provides information related to Scientific names, family, group and common names, as well as habitat, status of invasion in Mexico, pathways of introduction and links to other specialised websites. Some of the higher risk species already have a direct link to the alert page. It is important to notice that these lists are constantly being updated, please refer to the main page (http://www.conabio.gob.mx/invasoras/index.php/Portada), under the section Novedades for information on updates.
Invasive species - mammals is available from: http://www.conabio.gob.mx/invasoras/index.php/Especies_invasoras_-_Mam%C3%ADferos [Accessed 30 July 2008]
Spanish:
La lista de especies del Sistema de informaci�n sobre especies invasoras de m�xico cuenta actualmente con informaci�n aceca de nombre cient�fico, familia, grupo y nombre com�n, as� como h�bitat, estado de la invasi�n en M�xico, rutas de introducci�n y ligas a otros sitios especializados. Algunas de las especies de mayor riesgo ya tienen una liga directa a la p�gina de alertas. Es importante resaltar que estas listas se encuentran en constante proceso de actualizaci�n, por favor consulte la portada (http://www.conabio.gob.mx/invasoras/index.php/Portada), en la secci�n novedades, para conocer los cambios.
Especies invasoras - Mam�feros is available from: http://www.conabio.gob.mx/invasoras/index.php/Especies_invasoras_-_Mam%C3%ADferos [Accessed 30 July 2008]
Summary: English:
The species list sheet for the Mexican information system on invasive species currently provides information related to Scientific names, family, group and common names, as well as habitat, status of invasion in Mexico, pathways of introduction and links to other specialised websites. Some of the higher risk species already have a direct link to the alert page. It is important to notice that these lists are constantly being updated, please refer to the main page (http://www.conabio.gob.mx/invasoras/index.php/Portada), under the section Novedades for information on updates.
Invasive species - mammals is available from: http://www.conabio.gob.mx/invasoras/index.php/Especies_invasoras_-_Mam%C3%ADferos [Accessed 30 July 2008]
Spanish:
La lista de especies del Sistema de informaci�n sobre especies invasoras de m�xico cuenta actualmente con informaci�n aceca de nombre cient�fico, familia, grupo y nombre com�n, as� como h�bitat, estado de la invasi�n en M�xico, rutas de introducci�n y ligas a otros sitios especializados. Algunas de las especies de mayor riesgo ya tienen una liga directa a la p�gina de alertas. Es importante resaltar que estas listas se encuentran en constante proceso de actualizaci�n, por favor consulte la portada (http://www.conabio.gob.mx/invasoras/index.php/Portada), en la secci�n novedades, para conocer los cambios.
Especies invasoras - Mam�feros is available from: http://www.conabio.gob.mx/invasoras/index.php/Especies_invasoras_-_Mam%C3%ADferos [Accessed 30 July 2008]
Cruz, J. B., and F. Cruz. 1996. Conservation of the dark-rumped petrel Pterodroma phaeopygia of the Galapagos Islands, 1982-1991. Bird Conservation International 6:23-32.
Drake, Sara J.; Weltzin, Jake F.; Parr, Patricia D., 2003. Assessment of non-native invasive plant species on the United States Department of Energy Oak Ridge National Environmental Research Park. Castanea. 68(1). March 2003. 15-30.
Gargominy, O., Bouchet, P., Pascal, M., Jaffre, T. and Tourneu, J. C. 1996. Consequences des introductions d'especes animals et vegetales sur la biodiversite en Nouvelle-Caledonie. Rev. Ecol. (Terre Vie) 51: 375-401.
Summary: Consequences to the biodiversity of New Caledonia of the introduction of plant and animal species.
Summary: Consequences to the biodiversity of New Caledonia of the introduction of plant and animal species.
Gauss, C.I., Dubey, J.P., Vidal, D., Ruiz, F., Vicente, J., Marco, I., Lavin, S., Gortazar, C., Almeria, S. 2005. Seroprevalence Of Toxoplasma Gondii In Wild Pigs (Sus Scrofa) From Spain, Veterinary Parasitology 131: abstract. [Accessed 23 February, 2006 from Agricultural Research Service (ARS) (online database)]
Griffen, J. 1977. Ecology of the feral pig on Hawaii Island. Elepaio 37: 140�142.
Hook, T. and Todd, P. 1992. Mouse eradication on Mana Island. In Veitch, C. R., Fitzgerald, M., Innes, J. and Murphy, E. (eds) Proceedings of the national predator management workshop. Threatened Species Occasional Publication 3. Department of Conservation, Wellington. 33pp.
Huenneke, L. F., and P. M. Vitousek. 1990. Seedling and clonal recruitment of the invasive tree Psidium cattleianum: Implications for management of native Hawaiian forests. Biological Conservation 53:199-212.
ITIS (Integrated Taxonomic Information System), 2005. Online Database Sus scrofa
Summary: An online database that provides taxonomic information, common names, synonyms and geographical jurisdiction of a species. In addition links are provided to retrieve biological records and collection information from the Global Biodiversity Information Facility (GBIF) Data Portal and bioscience articles from BioOne journals.
Available from: http://www.cbif.gc.ca/pls/itisca/taxastep?king=every&p_action=containing&taxa=Sus+scrofa&p_format=&p_ifx=plglt&p_lang= [Accessed March 2005]
Summary: An online database that provides taxonomic information, common names, synonyms and geographical jurisdiction of a species. In addition links are provided to retrieve biological records and collection information from the Global Biodiversity Information Facility (GBIF) Data Portal and bioscience articles from BioOne journals.
Available from: http://www.cbif.gc.ca/pls/itisca/taxastep?king=every&p_action=containing&taxa=Sus+scrofa&p_format=&p_ifx=plglt&p_lang= [Accessed March 2005]
Jaksic, Fabian & Walton, Agustín & Jiménez, Jaime E. & Martinez, David R.. (2002). Invaders without frontiers: Cross-border invasions of exotic mammals. Biological Invasions. 4. 10.1023/A:1020576709964.
Kalesnik, Fabio; Acenolaza, Pablo, 2008. Regional distribution of native and exotic species in levees of the lower delta of the Parana river. Acta Scientiarum Biological Sciences. 30(4). OCT-DEC 2008. 391-402.
Kirk, T. 1896. The displacement of species in New Zealand. Transactions of the New Zealand Institute 28: 1�27.
Loewenstein, Nancy J.; Loewenstein, Edward F., 2005. Non-native plants in the understory of riparian forests across a land use gradient in the Southeast. Urban Ecosystems. 8(1). MAR 2005. 79-91.
Lorvelec, O. & Pascal, M. 2006. Les vert�br�s de Clipperton soumis � un si�cle et demi de bouleversements �cologiques. Revue d Ecologie (La terre et la Vie), 61, 2
Louette M. 1999. La Faune terrestre de Mayotte - Mus�e Royal de l Afrique Centrale, 247 p.
Summary: Synth�se g�n�rale sur la faune terrestre de Mayotte
Summary: Synth�se g�n�rale sur la faune terrestre de Mayotte
McEwan, Ryan W.; Rieske, Lynne K.; Arthur, Mary A., 2009. Potential interactions between invasive woody shrubs and the gypsy moth (Lymantria dispar), an invasive insect herbivore. Biological Invasions. 11(4). APR 2009. 1053-1058.
Merriam, Robert W., 2003. The abundance, distribution and edge associations of six non-indigenous, harmful plants across North Carolina. Journal of the Torrey Botanical Society. 130(4). October-December 2003. 283-291.
Merriam, Robert W.; Feil, Elisabeth, 2002. The potential impact of an introduced shrub on native plant diversity and forest regeneration. Biological Invasions. 4(4). 2002. 369-373.
Morris, Lorna L.; Walck, Jeffrey L.; Hidayati, Siti N., 2002. Growth and reproduction of the invasive Ligustrum sinense and native Forestiera ligustrina (Oleaceae): Implications for the invasion and persistence of a nonnative shrub.International Journal of Plant Sciences. 163(6). November 2002. 1001-1010.
Mus�um national d Histoire naturelle [Ed]. 2003-2006 . Sus scrofa. Inventaire national du Patrimoine nature
Summary: Available from: http://inpn.mnhn.fr/isb/servlet/ISBServlet?action=Espece&typeAction=10&pageReturn=ficheEspeceDescription.jsp&numero_taxon=60981[Accessed 20 March 2008]
Summary: Available from: http://inpn.mnhn.fr/isb/servlet/ISBServlet?action=Espece&typeAction=10&pageReturn=ficheEspeceDescription.jsp&numero_taxon=60981[Accessed 20 March 2008]
Osland, Michael J.; Pahl, James W.; Richardson, Curtis J., 2009. Native Bamboo [Arundinaria gigantea (Walter) Muhl., Poaceae] Establishment and Growth after the Removal of an Invasive Non-Native Shrub (Ligustrum sinense Lour., Oleaceae): Implications for Restoration. Castanea. 74(3). SEP 2009. 247-258.
Pascal, M., Barr�, N., De Garine-Wichatitsky, Lorvelec, O., Fr�tey, T., Brescia, F., Jourdan, H. 2006. Les peuplements n�o-cal�doniens de vert�b�br�s : invasions, disparitions. Pp 111-162, in M.-L. Beauvais et al., : Les esp�ces envahissantes dans l�archipel n�o-cal�donien, Paris, IRD �ditions, 260 p.+ c�d�rom
Summary: Synth�se des introductions d esp�ces de vert�br�s en Nouvelle-Cal�donie et �valuation de leurs impacts.
Summary: Synth�se des introductions d esp�ces de vert�br�s en Nouvelle-Cal�donie et �valuation de leurs impacts.
Probst J.-M. 1997. Animaux de la R�union. Azal�es Editions. 168 pp.
Rudge, M. R. 1976. A note on the food of feral pigs (Sus scrofa) of Auckland Island. Proceedings of the New Zealand Ecological Society 23: 83�84.
Spatz, G. and D. Mueller-Dombois. 1975. Succession patterns after pig digging in grassland communities on Mauna Loa, Hawaii. Phytocoecologia 3:346-373.
Stone, C. P., L. W. Pratt, and J. T. Tunison. 1992. Responses of Hawaiian ecosystems to removal of feral pigs and goats. Pages 666-704 in C. P. Stone, C. W. Smith, and J. T. Tunison, editors. Alien plant invasions in native ecosystems of Hawai i: management and research. Cooperative National Park Resources Studies Unit. University of Hawaii at Manoa., Honolulu, HI.
Stromayer, Karl A. K.; Warren, Robert J.; Harrington, Timothy B., 1998b. Managing Chinese privet for white-tailed deer. Southern Journal of Applied Forestry. 22(4). Nov., 1998. 227-230.
Stromayer, Karl A. K.; Warren, Robert J.; Johnson, A. Sydney; Hale, Philip E.; Rogers, Carolyn L.; Tucker, Christopher L., 1998a. Chinese privet and the feeding ecology of white-tailed deer: The role of an exotic plant. Journal of Wildlife Management. 62(4). Oct., 1998. 1321-1329.
Taylor, Constance E. S.; Magrath, Lawrence K.; Folley, Patricia; Buck, Paul; Carpenter, Sydney, 1996. Oklahoma vascular plants: Additions and distributional comments. Proceedings of the Oklahoma Academy of Science. 76(0). 1996. 31-34.
Tisdell, C. A. 1982. Wild Pigs: Environmental Pest or Economic Resource? Pergamon Press, Sydney.
USDA, ARS, 2010. Taxon: Ligustrum sinense Lour. National Genetic Resources Program. Germplasm Resources Information Network - (GRIN) [Online Database]. National Germplasm Resources Laboratory, Beltsville, Maryland.
Summary: Available from: http://www.ars-grin.gov/cgi-bin/npgs/html/taxon.pl?22095 [Accessed 10 March 2010]
Summary: Available from: http://www.ars-grin.gov/cgi-bin/npgs/html/taxon.pl?22095 [Accessed 10 March 2010]
USDA, NRCS. 2010. Ligustrum sinense Lour. Chinese privet. The PLANTS Database (http://plants.usda.gov, 10 March 2010). National Plant Data Center, Baton Rouge, LA 70874-4490 USA.
Summary: Available from: http://plants.usda.gov/java/profile?symbol=LISI [Accessed 10 March 2010]
Summary: Available from: http://plants.usda.gov/java/profile?symbol=LISI [Accessed 10 March 2010]
Wilcox, Joshua; Beck, Christopher W., 2007. Effects of Ligustrum sinense Lour. (Chinese privet) on abundance and diversity of songbirds and native plants in a southeastern nature preserve. Southeastern Naturalist. 6(3). 2007. 535-550.
Contact
The following 3 contacts offer information an advice on Sus scrofa
de Garine-Wichatitsky,
Michel
Michel
Geographic region: Pacific, southern Africa
Ecosystem: Terrestrial
Ecosystem: Terrestrial
Organization:
Centre de coop�ration internationale en recherche agronomique pour le d�veloppement
Address:
CIRAD/UR22, Integrated Wildlife Management. 37 Arcturus Road, Highlands, P.O. Box 1378, Harare, Zimbabwe
Phone:
(263)4 443422
Fax:
Hadjisterkotis,
Eleftherios
Eleftherios
Organization:
Ministry of the Interior
Email:
Address:
1453 Nicosia, Cyprus
Phone:
Fax:
McIlroy,
John
John
Organization:
Email:
Address:
38 Hempleman Drive, Akaroa, 8161, New Zealand
Phone:
+64 3 3047210
Fax:
+64 3 304 8430
Sus scrofa
pig, Wildschwein, razorback, te poaka, kuhukuhu, poretere, petapeta, kune-kune
pig, Wildschwein, razorback, te poaka, kuhukuhu, poretere, petapeta, kune-kune
Date assessed
Year published
Eicat category
Justification for EICAT assessment
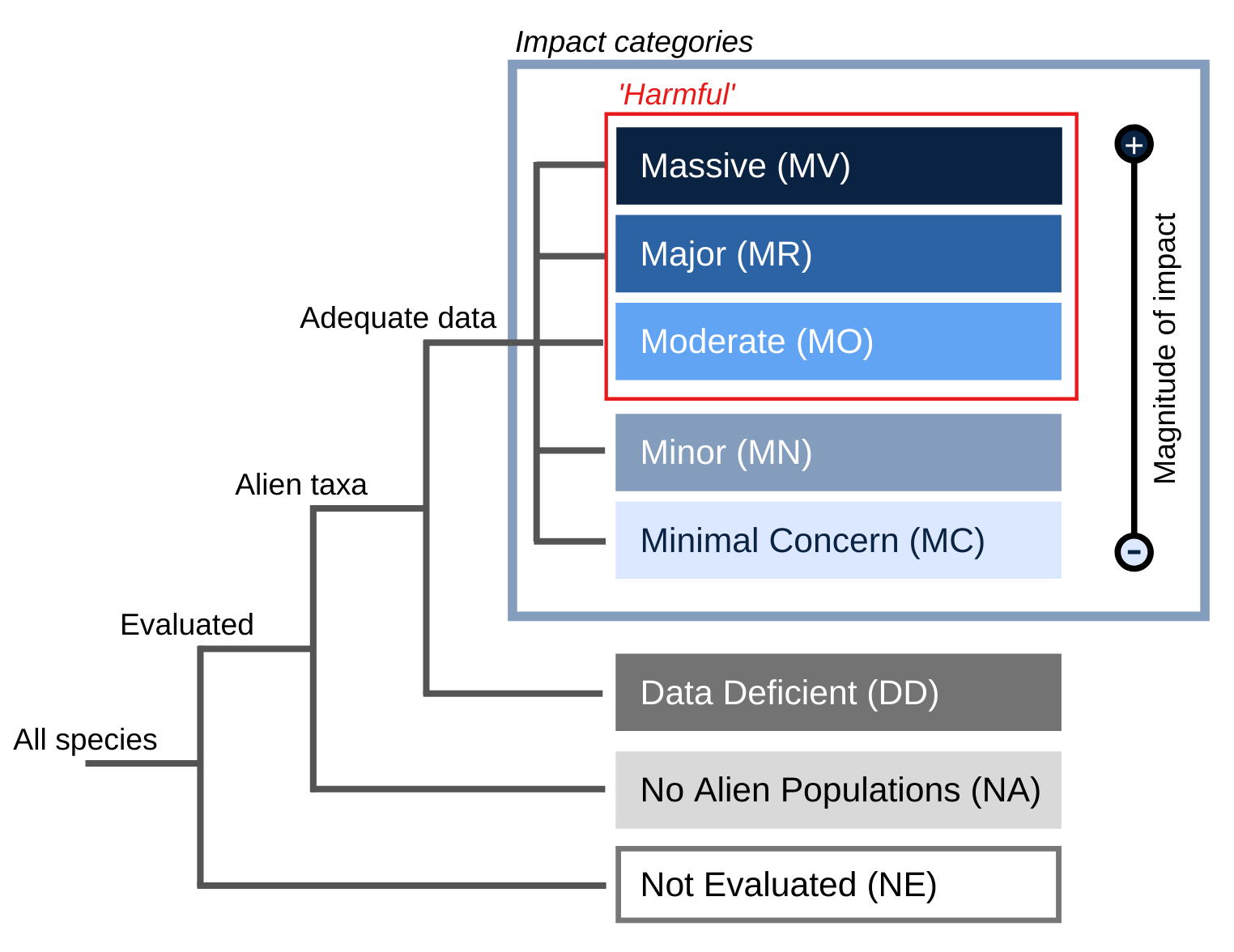
Confidence rating
Mechanism(s) of maximum impact
Countries of most severe impact
Description of impacts
Assessor
Contributors
Reviewers
Recommended citation
(2026). Sus scrofa. IUCN Environmental Impact Classification for Alien Taxa (EICAT).